地球上に現存している植物は 30 万種以上である。植物は動物と異なり移動ができないため、常に光不足、乾燥や捕食などのストレスにさらされ、独自の形態やストレス耐性を獲得してきた。植物科学は、こうした植物の形態や植物の生理現象などを取り扱う学問である。このページでは植物化学に関連する用語とその説明を掲載している。
原形質連絡 (plasmodesmata)
隣り合う細胞の原形質間に存在する物質輸送や情報交換用の通路である。通路はデスモ小管とその外側を覆う細胞膜からなる。デスモ小管の表面にはアクチンフィラメントなどのタンパク質が存在し物質輸送に働く。細胞壁で保護されている植物細胞にとって、原形質連絡は細胞間の活動に不可欠である。
原形質分離 (plasmolysis)
植物細胞を高張液に浸すと、細胞内にある水が細胞外に流出し、細胞膜に覆われた原形質の部分が収縮する。一方、細胞壁はほとんど変形しないために、収縮した細胞膜とその内部の原形質が細胞壁から分離する。これを原形質分離という。動物細胞には細胞壁が存在しないため、原形質分離が起こらない。
原形質流動 (cytoplasmic streaming)
生きている細胞内部で、細胞小器官などの原形質が、細胞骨格とモータータンパク質などの相互作用により流れ動くこと。
窒素固定 (nitrogen fixation)
空気中に存在する窒素分子を取り込み、アンモニアや硝酸塩などの反応性の高い無機窒素化合物に変更すること。根粒菌、アゾトバクター、クロストリジウム、藍藻類やメタン菌などによって行われる。窒素固定は ATP の消費を伴う。
窒素同化 (nitrogen assimilation)
遊離窒素や無機窒素化合物などを生体内に取り入れて、有機窒素化合物を合成すること。植物では植物体に取り込まれたアンモニアイオンをグルタミン酸に結合させ、グルタミン、α-ケトグルタル酸などの有機化合物に次々と受け渡し、最後にアンモニアイオンを各種の有機酸に転置させて、多様なアミノ酸を生成する。
脱窒 (denitrification)
窒素化合物の窒素を遊離窒素に変換し外界に放出すること。主に脱窒菌の働きによって硝酸イオンが窒素分子あるいは酸化窒素に変換される。脱窒菌は脱窒素作用により ATP を合成する。
植物ホルモン (plant hormone)
植物体により生産され、個体内を移動し他の場所にて生理過程を調節する物質のこと。低濃度で作用する。
| オーキシン auxin |
屈光性の研究などにより最初に発見され、インドール酢酸-3-酢酸(IAA)などの化合物として天然に存在する。主に、植物の伸長成長に関わる。また、オーキシンは屈光性と屈地性などにも関わる。
【促進】茎・根の伸長成長、細胞分裂、組織文化、発根、子房の成長(果実の肥大、単為結実) 【抑制】側芽の成長、落葉・落果 |
| ジベレリン gibberellin |
細胞の伸長促進、種子の発芽促進や休眠打破の促進に関わるホルモンとして知られる。馬鹿苗病から発見され、現在では 70 種類以上見つかっている。また、単為結実を促進するため種なしブドウの生産などにも用いられる。
【促進】伸長成長、休眠打破、種子の発芽、花芽形成、開花、単為結実 【抑制】 |
| サイトカイニン cytokinin |
オーキシンとともに細胞分裂、シュート形成の誘導効果をもつ化合物一群の総称。主に根で合成され、道管を通って地上部に輸送される。分裂組織、未熟な種子、形成途中の維管束などで濃度が高い。
【促進】細胞分裂(オーキシン存在下)、シュート形成(オーキシン存在下)、側芽の成長 【抑制】老化 |
| アブシジン酸 abscisic acid |
乾燥などのストレスに応答して合成されることからストレスホルモンとして知られる。非メバロン酸経路により合成され、植物体全体にわたって広く分布する。また、種子の発達過程において種子中に蓄積され、種子の成熟、休眠を促進する。
【促進】気孔の閉鎖、種子の成熟、種子休眠、芽の休眠、落果・落葉の促進 【抑制】 |
| エチレン (ethene) |
果実の成熟に関与する気体の植物ホルモンである。病原菌の感染や組織が障害を受けた時にも生成され、防御応答を誘導する。リンゴ、アボカドなどは特に多くエチレンを放出する。
【促進】発芽、開花、果実の成熟、落葉の離脱、老化 【抑制】細胞分裂 |
| フロリゲン florigen |
花芽形成を誘導する植物ホルモン様物質。 |
| ジャスモン酸 jasmonic acid |
植物ホルモン様物質。果実の熟化、老化促進、休眠打破などを誘導する。また、障害などのストレスに応答して合成されることから耐性誘導ホルモンとしても知られる。 |
重複受精 (double fertilization)
被子植物に特有な受精様式で、2つの精核(精細胞)が卵細胞と中央細胞のそれぞれと受精する現象をいう。被子植物の受粉では、花粉から2個の精核(n)が胚嚢に送り込まれ、その内の一つは卵細胞(n)と受精し、胚(2n)となる。これを生殖受精という。もう一つ精核は極核(2n)と合体し、胚乳(3n)となる。これを栄養受精という。
種子休眠 (seed dormancy)
種子が発芽に適した条件下においても発芽しないことをいう。種子が、発芽の必須条件が与えられても発芽しないのは、次のようなことが考えられる。
- 固い種皮が、水分や酸素がなどを通さないために、発芽に至らない。このような種子では、種皮に傷が付くことによって休眠が打破される。(クローバー、オナモミなど)
- 種子の胚の発達が不十分で未熟なために発芽に至らない。このような種子は、一定期間経過すると種子が成熟するため、休眠が打破される。(イチョウなど)
- 発芽抑制物質により、種子が発芽に至らない。このような種子は、種皮が薄く、胚も発達していながら休眠が維持される。休眠を打破するには、種子を低温にさらしたり、水に浸漬したりするなどの処理が必要。(リンゴ、イネなど)
休眠打破 (breaking dormancy)
休眠状態の種子が胚の発生・成長を再開することをいう。休眠打破の引き金となる要因は様々ある。例えばクローバーやオナモミなどの固い種皮を持つ種子の場合は、種皮に傷を付けることで休眠が打破される。イチョウの種子などのように、一定期間が経過すると休眠が打破される。また、イネなどのように、水に浸漬することで休眠が打破される。
光補償点 (light compensation point)
植物は光の存在下で呼吸と光合成を同時に行っている。呼吸による二酸化炭素の放出量と光合成による二酸化炭素吸収量が釣り合い、見かけ上、二酸化炭素の出入りがなくなるときの光の強度を光補償点という。強い光の下で育った陽葉の光補償点は高く、逆に弱い光の下で育った陰葉の光補償点は低い。
光飽和点 (light saturation point)
光合成速度は一般的に光の強さ(照度)が強くなるほど上昇する。しかし、ある一定の照度に達すると、光合成速度が上昇しなくなる。このときの光の照度を光飽和点という。このとき、光合成の制限要因は二酸化炭素あるいは温度となる場合がある。
C4 植物 (C4 plant)
光合成の過程でカルビン-ベンソン回路の他に、二酸化炭素を濃縮するための C4 回路(ハッチ-スラック回路)の両方を持つ植物をいう。高温や乾燥などの悪条件では、二酸化炭素の取り込み効率が悪くなる。そのため、カルビン-ベンソン回路のみで行わる光合成反応は、低二酸化炭素濃度のために光呼吸が生じる。このような状況において、C4 植物は二酸化炭素をいったん維管束鞘細胞に取り込み、C4 回路により有機酸に固定する。次に、維管束鞘細胞で固定・濃縮された二酸化炭素を、カルビン-ベンソン回路に与えて光合成を行う。このように C4 植物は高温や乾燥などの条件下においても高濃度な二酸化炭素を利用できるため、光呼吸を回避することができる。トウモロコシ、サトウキビなどが C4 植物である。
CAM 植物 (CAM plant)
CAM 植物はカルビン-ベンソン回路と CAM 回路の両方を持つ植物である。CAM 植物は夜に気孔を開けて、CAM 回路により二酸化炭素の取り込みを行う。昼間に CAM 回路により固定した二酸化炭素をカルビン-ベンソン回路に渡し、光合成を行う。こうした CAM 植物は、砂漠などの過酷な環境下でも、水分の損失を最小限に抑えながら光合成できる。サボテンやベンケイソウなどが CAM 植物である。
蒸散 (transpiration)
植物の地上部から大気中へ水蒸気が放出される現象である。蒸散の大部分は気孔の集中している葉の裏側で起こる。その他に、茎化や葉の表側や果実においても蒸散が確認される。
リブロース1,5-ビスリン酸カルボキシラーゼ、RubisCO (ribulose 1,5-bisphosphate carboxylase)
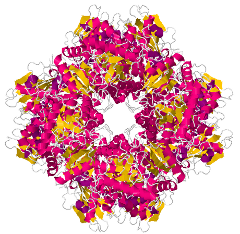
RubisCO は大サブユニット 8 個および小サブユニット 8 個からなるヘテロ 16 量体のタンパク質であり、活性中心にはマグネシウムイオンが配置されている。カルビン-ベンソン回路に存在し光合成に関わる。カルボキシラーゼ活性とオキシゲナーゼ活性の両方を持つ。
細胞内の二酸化炭素濃度が高いとき、Rubisco はカルボキシラーゼ活性を示す。1 分子のリブロース1,5-ビスリン酸(RuBP)に 1 分子の二酸化炭素を固定し、2 分子の 3-ホスホグリセリン酸(PGA)を生成する反応を触媒する。
一方、、細胞内の酸素濃度が高いとき、RubisCO はオキシゲナーゼ活性を示し、上記と異なる反応を触媒する。この反応は、RuBP が酸化され、3-ホスホグリコール酸と 3-ホスホグリセリン酸が生成される。3-ホスホグリコール酸は細胞内で代謝され最終的に二酸化炭素として放出される。このように、光照射下において二酸化炭素を放出する反応を光呼吸とよぶ。
光呼吸 (photorespiration)
植物が光照射下において二酸化炭素を放出する反応を光呼吸とよぶ。光合成において、二酸化炭素を有機酸に固定する反応を触媒する酵素は RubisCO である。この反応は、カルビン-ベンソン回路で行われ、1 分子のリブロース1,5-ビスリン酸(RuBP)に 1 分子の二酸化炭素を固定し、2 分子の 3-ホスホグリセリン酸(PGA)を生成する。
しかし、温度が上昇すると細胞内の二酸化炭素と酸素の溶解度が低下する。特に、二酸化炭素の溶解度の低下が激しいため、溶存酸素の濃度が相対的に大きくなる。このとき、RubisCO は RuBP を酸化し、3-ホスホグリコール酸と 3-ホスホグリセリン酸を生成する反応を触媒するようになる。3-ホスホグリコール酸は細胞内で代謝され二酸化炭素として放出する。この反応は ATP と NADPH の消費を伴う。
二酸化炭素の量が光エネルギーに比べ不十分であるとき、過剰な光エネルギーによる障害を防ぐなどの生理機能を果たしていると考えられる。また、C4 植物や CAM 植物は二酸化炭素を濃縮する経路を持つため、光呼吸がほとんど起こらないと考えられる。
頂芽優勢 (apical dominance)
植物の茎の先端にある頂芽の伸張成長が促進され、側芽の成長が抑制されること。オーキシンなどの植物ホルモンが関係する。
光形態形成 (photomorphogenesis)
植物が光を受容することによって、生長や分化などの形態が制御されること。光受容体としてフィトクロム(赤色光・遠赤色光)、クリプトクロム(青色光)やフォトトロピン(青色光)が知られている。
| フィトクロム phytochrome |
フィトクロムは赤色光吸収型(Pr 型)と遠赤色光吸収型(Pfr 型)が存在し、両者の間を可逆的に変化する。例えば、レタスの種子に対して赤色光を照射したとき、フィトクロムは Pr 型となり赤色光を吸収し、種子の発芽を誘導する。しかし、遠赤色光を照射したとき、フィトクロムは Pfr 型に変化し遠赤色光を吸収するようになり、種子の発芽を阻害する。植物のみに存在する。 |
| クリプトクロム cryptochrome |
青色光を受容する光受容体。花芽形成、伸長、概日リズムなどの調節に関わる。緑藻から高等植物などに存在する。動物にはクリプトクロムに似たタンパク質が存在する。 |
| フォトトロピン phototropin |
青色光を受容する光受容体。光屈性、気孔の開閉などの調節に関わる。植物のみに存在する。 |
種子繁殖 (seed propagation)
種子によって繁栄することを種子繁殖という。種子は一般に受精によって形成されるが、受精しないで種子形成する無配偶生殖もある。被子植物は雄ずいと雌ずいが一つの花の中にある両生花を持つことが多く、自家受粉しやすい構造となっている。特に自家受粉により繁栄する植物のことを自殖性植物という。
栄養繁殖 (vegetative propagation)
ジャガイモ、いちご、ぶどう、百合などのように地下茎や根、むかごなどから増殖することを栄養繁殖という。ぶどうなどのように種をつける植物も存在するが、種子繁殖する植物に比べ効率が悪い。
ABC モデル (ABC model)
花の形態形成に関わる MADSbox 遺伝子は、機能的に 3 種類に分けることができる。それぞれ、A、B、C の遺伝子と例えられる。A 遺伝子が単独で発現する部位はがくが形成される。C 遺伝子が単独で発現する部位は雌ずいが形成される。これに加え、A と B 遺伝子が同時に発現する部位は花弁が形成され、B と C 遺伝子が同時に発現する部位は雄ずいが形成される。また、A 遺伝子と C 遺伝子は拮抗しているため、A 遺伝子が欠損するとそこに C 遺伝子が発現する。このような花の形態形成モデルを ABC モデルという。
花粉 (pollen)
花粉は葯から作られる。葯は胞原細胞を含み、発達の過程において胞原細胞は側壁細胞と胞子形成細胞に分化する。胞子形成細胞が発達すると花粉母細胞ある。一つの花粉母細胞が減数分裂し、四分子とよばれる 4 つの半数性細胞ができる。この半数性細胞が休止期を経て再び細胞分裂を行い、栄養細胞と雄原細胞を形成する。雄原細胞は色素体とミトコンドリアを持ち、さらに細胞分裂により 2 つの精細胞を形成する。
雄原細胞が精細胞になる時期は植物種によって異なり、イネやアブラナなどでは葯の中ですでに細胞分裂を開始するが、ナスやマメなどでは受粉後、花粉管中で細胞分裂を行う。
重複受精 (double fertilization)
成熟した胚嚢は、合点側に 3 つの反足細胞、中央に大きな中央細胞、珠孔側に 2 つの助細胞と 1 つの卵細胞を持つ。中央細胞は 2 つの極核を持つ。受精は、2 つの精細胞が花粉管を通り珠孔から胚珠に侵入し、1 つの精細胞が卵細胞と核融合し 2n の受精卵となる。残りのもう 1 つの精細胞は中央細胞の極核と核融合し 3n の胚乳になる。このような被子植物に特有な受精方法を重複受精という。
不和合性 (incompatibiligy)
花粉と雌ずいが正常で、それぞれに生殖能力を有しているにもかかわらず、受粉しても受精が起こらないことを不和合性という。自家受粉時に示す不和合性をとくに自家不和合性といい、また種間受粉時に示す不和合性をとくに種間不和合性という。