Cardamine (タネツケバナ)属は Arabidopsis thaliana の近縁種であり、およそ 150 以上の種が多様な環境に適応して生息していると言われている。タネツケバナ属の種はゲノムサイズが小さく、また、その多様性のゆえに、植物生態学の研究によく用いられる。また、タネツケバナ属にはいくつかの異質倍数体が存在し、その親種に明確な形質の違いが見られることから、タネツケバナ属の植物は異質倍数体の研究対象ともなっている。
生息環境
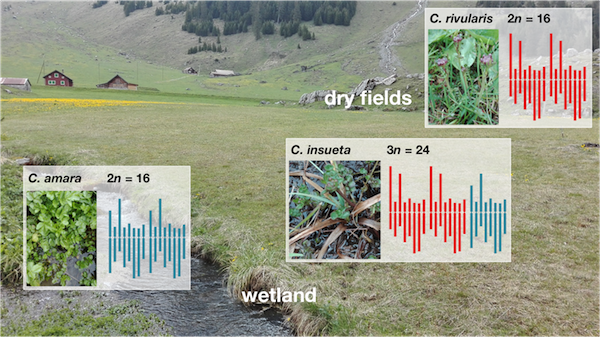
タネツケバナ属の植物の研究は、スイスアルプスの Urnerboden 地域をモデルとしているものが多い。Urnerboden 地域はアルプスの山々に囲まれた地域である。春先になると、アルプス山脈からの雪解け水が山の根元に流れ込み小川が形成される。5 月末から 6 月にかけて気温が上昇すると、雪解けが早くなり、小川が溢れて近くの草地までが水浸し状態になる。しかし、小川からやや遠く離れた場所までは冠水されることはない。C. amara (2n=2x=16, AA) は Urnerboden 地域において、小川沿いあるいは水中で生息している。また、C. rivularis (2n=2x=16, RR) はほとんど冠水されない場所に分布している。両者の交雑種(異質倍数体)である C. insueta (2n=3x=24, RRA) は小川沿いから冠水されるような場所に広く分布している(Sun et al., 2020)。C. rivularis が冠水にさらされると生存できなくなることが知られているため、C. insueta が川沿いであるいは一時冠水されても生存できるようになったのは、おそらく C. amara から何らかの冠水ストレスに耐性を持つ遺伝子を受け継いで、冠水ストレス耐性を持つようになったのではないかと考えられる。
異質倍数体
Urnerboden 地域はかつて森林地域であったが、19 世紀末の農地開墾によって森林が消滅した。それまでは、森林によって生殖隔離され、C. amara は川沿いで生息し、C. rivularis は比較的乾いた環境に生息していた。しかし、森林が消滅することにより、C. amara と C. rivularis の生息域が拡大し、両者の生息域が重なるようになった。両者の生息域が重なる場所において、異種間交雑が行なわれ C. insueta などの新種が誕生したと言われている(Mandáková et al., 2013)。
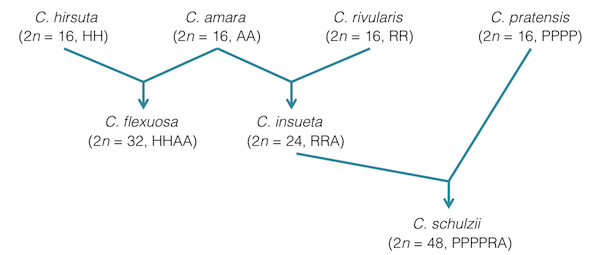
タネツケバナ属の 2 倍体には C. hirsuta、C. rivularis、C. amara などが知られている。これらが異種間交雑することによって、C. flexuosa、C. insueta や C. schulzii などの異質倍数体が誕生したと言われている(Urbanska, 1981, Mandáková et al., 2013, Zozomová-Lihová et al., 2014)。
繁殖様式
Uernerboden 地域で確認された C. amara と C. insueta は種子繁殖とクローン繁殖の両方が可能である。クローン繁殖について、C. amara はランナーにより、C. insueta は栄養分体(plantlet)によるものである。また、両者がハイブリッド種は C. rivularis はクローン繁殖のみで、ランナーを形成するが、主に plantlet によって繁殖している(Urbanska, 1981)。
Uernerboden 地域では春が過ぎてから、牛などがこの場所を通るために、これらの植物は種子を作る前に、牧草とともに食べられることが多い。そのため、Urnerboden 地域においては、種子による繁殖よりもクローン繁殖の方が効率的であると考えられる。
C. amara
C. amara は種子繁殖とクローン繁殖の両方が可能である。C. amara の種子の発芽率が高く、種子繁殖がよく行なわれている。その一方で、クローン繁殖もさんかに行なわれている。C. amara のクローン繁殖は、secondary rosettes と開花しているシュートの頂端からランナーを形成する形態をとっている。C. amara は 2 つの繁殖様式をバランス良く利用してい繁殖していると考えられている。しかし、水辺などでは、クローン繁殖により形成された集落が確認されている。これらの集落で生息している個体は花を咲かせるが、多くの場合、自家受粉してしまうために種子ができない。
Uerner Boden 谷で撮影した、小川の側で生息している C. amara の個体。

Uerner Boden 谷で撮影した、小川の中でクローン繁殖によって形成した C. amara のコロニー。C. amara の 1 個体のサイズが大きいので、2 倍体ではないと考えられる。

C. rivularis
C. rivularis は主に種子繁殖により繁殖している。しかし、クローン繁殖により繁殖することも確認されている。C. rivularis のクローン繁殖は、葉と茎の付け根から plantlet を発生させる形態をとっている。plantlet が発生するタイミングについてはわかっていない。

C. insueta
C. insueta は C. amara と C. rivularis が自然交雑することによって誕生した種である。C. insueta はC. amara由来のAゲノムを1本と C. rivularis 由来のRゲノムを2本持つ異質 3 倍体である。C. insueta はゲノムセットを奇数個に持つため、ほとんどの場合、減数分裂が失敗する。そのため、C. insueta は、開花するが種子が形成されない。
C. insueta は主にクローン繁殖によって繁殖している。その繁殖形態は非常は複雑で、C. amara と C. rivularis の両方の親種に由来する形質を持つ。すなわち、C. insueta は C. amara のように secondary rosettes からランナーを形成して、繁殖することができるだけでなく、C. rivularis のように葉と茎の付け根から plantlet を形成して繁殖することもできる。C. insueta は主に plantlet によるクローン繁殖を行っている。とくに、草刈りが行われた直後に、C. insueta の個体の多くは刈り取られ、折れたシュートが地面に落ちる。地面に着いたシュートは plantlet を発生させて成長するようになる。そのため、草刈のあとには、C. insueta の生息密度が非常に高いことがわかあっている。
C. insueta の個体サイズは C. rivularis より少しだけ大きいようにみえる。C. amara に比べれば明らかに小さい。茎の太さは、C. insueta と C. rivularis とほぼ同じぐらい細い。また、C. insueta の葉の形は 2 種類の形態があり、茎の根元の部分にあるロゼットは C. amara の葉の形に似て、その一方で、そのほかの部位にある葉の形は C. rivularis の葉の形に似ている。
Uerner Boden 谷で撮影した、小川のそば C. insueta の個体。


References
- A Recently Formed Triploid Cardamine insueta Inherits Leaf Vivipary and Submergence Tolerance Traits of Parents. Front. Genet. 2020, 11:567262. DOI: 10.3389/fgene.2020.567262
- Reproductive Strategies in some Perennial Angiosperms Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich. 1981, 126(4):269-84.
- The More the Merrier: Recent Hybridization and Polyploidy in Cardamine. Plant Cell. 2013, 25(9):3280-95. DOI: 10.1105/tpc.113.114405
- Multiple hybridization events in Cardamine (Brassicaceae) during the last 150 years: revisiting a textbook example of neoallopolyploidy. Ann Bot. 2014, 113(5):817-30. DOI: 10.1093/aob/mcu012